



出品| 新浪科技《科学大家》、知识分子
撰文| 饶毅 生命科学家、北京大学教授
涉及人类生命的技术进步是把双刃剑。2012年诞生的简便而强大的基因编辑技术(CRISPR/Cas9)是分子生物学重大技术进步,合理应用可增强人类福祉,不当应用可给公共安全和人类社会带来危机。
基因编辑人类体细胞与基因编辑生殖细胞有着本质的不同,体细胞编辑只影响个人而不带来新的伦理问题;而生殖细胞经过受精、胚胎发育、出生、长大成人结婚后将与其他人产生后代,会遗传给他人,编辑的后果将扩散,进而影响人类社会。体细胞的基因编辑可能治疗人类很多疾病,它与现有框架已接受的基因治疗类似,可按现有法律法规和监管办法,在个体安全性、作用有效性和方法稳定性确定后使用,但针对人类生殖细胞的基因编辑则超出现有伦理框架,它的特征是医学必要性低、技术可控性低、群体危害性高。
绝大多数携带疾病罹患基因的家庭,可用无伦理争议的筛查技术而无需生殖细胞基因编辑。一旦允许人类生殖细胞基因编辑,就难以限定可以编辑什么基因、编辑多少基因、由谁编辑基因等。中国需要立即调查处理已出现违反政策、规定、伦理的人类生殖细胞基因编辑问题,尽快加强监管弥补漏洞,出台有针对性的措施确保基因编辑技术不被滥用,制定人类基因编辑的法律法规:允许体细胞基因编辑,禁止人类生殖细胞的基因编辑。
缺乏监管的人类生殖细胞基因编辑可能带来对人类社会安全危害,相关科技人员和医生必须严格遵守法律、法规和伦理规范。缺乏法律法规和伦理规范可能带来国家之间的基因竞赛,因此需要国际社会协调制定国际公约,并将国际公约转化为各国的法律,以保护全人类共同利益。
深刻认识基因是人类智慧绚丽的结晶、合规操作基因是增进人类福祉可能的途径。但科学技术的进步有时是把双刃剑,特别是涉及人类生活和生命的时候。如何规范基因编辑是今天人类社会面临的严峻考验之一。

2012年,新的基因编辑技术CRISPR/Cas9诞生。这一新的技术作用强大,而且简单易用,因此门槛低、影响特别大。在什么范围应用如此强大的基因编辑技术,成为科学界、社会、国家乃至人类面临的巨大问题。对一般体细胞进行基因编辑,可在现有法律框架下进行监管问题,符合伦理规范不影响人类遗传、不扩散到他人。而对人类生殖细胞进行基因编辑,则超出患者个人治疗、预防和健康范畴,会对人类社会遗传带来不可估量的影响,挑战现有科研规范、有违人类道德伦理范围,也是国内外有关部门(如科技部、卫健委、中国科学院、中国工程院、中国医学科学院,美国国立健康研究院等)、科学学会和团体(中国遗传学会、细胞生物学会等)对之普遍谴责的原因。社会上对缺乏透明和监管的基因编辑感到焦虑和不安。
这是自2015年中国广州研究者第一次基因编辑人类生殖细胞后,法律法规没有跟上技术发展的必然发展。基因编辑人类生殖细胞具有“两低一高”的特点:医学必要性低、技术可控性低、群体危害性高。绝大多数病人家庭无需基因编辑、而通过基因测序后进行受精卵的选择就可以做到,且副作用更小。
基因编辑技术非常容易,所以很多人能够做,学生物的本科生就可能到县医院做。而选择改变什么性状、改变什么基因以及多少基因很难取得共识,由此会导致一片混乱。一旦允许人类生殖细胞基因编辑,就很难阻止大规模地改变人类基因的问题,而且少数人决定的基因编辑后果将会扩散到整个人群,这样就会带来整个人类的灾难。突发事件已经表明,对于基因编辑这种技术的使用,仅仅依赖科研人员的自律是不够的,必须制定相关的法律法规予以规范,否则基因编辑技术滥用的后果是科技工作者个人、国家乃至人类都难以承受的。
现在国内外讨论大多集中于基因编辑产出的婴儿个人安全问题,包括黑箱操作、质量低劣、缺乏基本的动物研究背景就极快速度用人做试验等,这些讨论其实漏洞百出。个人安全固然重要,但还有更大的问题:改变体细胞的基因是个人和家庭的问题,而改变生殖细胞基因是群体和人类的问题,前者可以由个人和家庭决定,后者需要国家和国际进行规范。

亡羊补牢,犹未为晚。中国应该以立法为目标进行严肃讨论,研究制定符合中国人民和人类利益、对科学界有约束力、对个人有威慑力以及切实可行的法律法规,从而对生殖细胞基因编辑进行规范管理。目前,我们还没有进行人类生殖细胞编辑的迫切理由,可禁止其临床实践,直到有医学刚需并解决了伦理和法律问题之后。
基因:科学和技术
自孟德尔发现遗传规律以来的150多年,经过遗传学、生物化学、微生物学和分子生物学的发展带来很多在科学和技术方面的进步。今天以基因为核心的分子生物学渗透了生物学、医学和农业科学等学科,而以基因分析和基因操纵为主的分子生物学技术也早已走出实验室被广泛应用于药学、农业和医学等领域。

百年基础研究,从遗传学到分子生物学
1866年,位于现今捷克布鲁恩修道院的孟德尔发表了他历经十年的研究结果,揭示遗传的基本规律,提出遗传因子的雏形(Mendel,1866)。1871年,瑞士科学家Miescher报道后来称之为“核酸”的化学物质(Miescher,1871),以后我们才知道DNA(脱氧核糖核酸)和RNA(核糖核酸)。1944年,美国洛克菲勒医学研究所Avery等提出DNA是遗传的物质基础(Avery, McCleod, McCarty, 1944)。1953年,美国遗传学家Watson和英国物理学家Crick提出DNA的双螺旋结构模型(Watson and Crick, 1953)。1953年至1965年,Crick及英国、美国多位科学家竞争、合作、交叉,完全解析了生物普遍适用的遗传密码,奠定了分子生物学的框架,促进了一个新学科的诞生和蓬勃发展。
1973年,美国旧金山加州大学的Boyer和斯坦福大学的Cohen发明现今常用的重组DNA技术(Cohen et al。, 1973),从此科学家可以剪切DNA,按需求拼接不同的DNA。1977年,英国剑桥大学Sanger等和美国哈佛大学Gilbert等分别发明DNA测序方法(Sanger et al。, 1977;Maxam and Gilbert, 1977)。这两项技术结合起来,不仅改变了分子生物学的进程,而且把科学带出了学术的象牙塔,应用于生物技术、医学、农业、药学等领域。1976年,Boyer与投资人Swanson建立了世界上第一个以DNA为基础的现代生物技术企业Genentech,象征着现代生物技术产业的诞生。
对于基因的操纵,科学家早期只能在细菌进行,如把人的胰岛素基因克隆到无致病性的细菌中,让后者为人类生产蛋白质药物。随后基因操作技术逐渐发展,可在其他物种中进行,如1987年美国科学家Capecchi等发明了小鼠基因敲除的方法(Thomas and Capecchil, 1987),对生物学的研究带来很大推动。
但在很长时间,科学家只能对少数生物进行基因操纵(如细菌、酵母、线虫、果蝇、斑马鱼、小鼠等),而不能对大多数生物进行基因操作。
在这样的背景下,一个新的技术诞生于意想不到的研究。1987年,日本大阪大学微生物学家中田久郎(Atsuo Nakata)实验室发现细菌中存在后来称为CRISPR的DNA序列(Ishino et al。, 1987),经过日本、荷兰、西班牙、立陶宛、美国、法国、英国等国多个相对默默无闻科学家的基础研究,发现CRISPR可与CAS蛋白质组成细菌的一种防御系统,可编辑剪切特定的DNA序列(Nakata et al。, 1989; Hermans et al。,1991; Groenen et al。,1993; Mojica et al。, 1993; Mojica et al。,1995; Mojica et al。, 2000; Jansen et al。,2002; Mojica et al。, 2005; Haft et al。,2005; Pourcel et al。,2005; Bolotin et al。, 2005; Makarova et al。,2006; Barrangou et al。,2007; Horvath et al。, 2008; Mojica et al。, 2009; Brouns et al。, 2008; Marraffini et al。, 2008; Deveau et al。, 2008; Garneau et al。, 2010; Hale et al。, 2009; Horvath and Barrangou; 2010; Deltcheva et al。,2011; Sapranauskas et al。, 2011; Hale et al。, 2012)。

基因编辑技术CRISPR/Cas9技术发现的三位先驱
2012年,法国科学家Emmanuelle Charpentiere和美国科学家Jennifer Doudna合作(Jinek et al。, 2012),与立陶宛的Siksnys实验室(Gasiunas et al。, 2012)分别证明可在体外重建CRISPR/Cas9体系,用于编辑特定的靶基因。这一方法被发明后,很快被用于各种细胞和各种生物,并在很多细胞和生物中证实非常有效。从此,我们改变DNA就不再局限于几种生物,特别是研究用的生物,而可以用于很多其他物种。其后几年,有关CRISPR/CAS9应用的论文如雨后春笋、层出不穷。这一技术也得到不断改进,使用更加方便,效率更高。
在这一过程中,海内外华人科学家,如美国MIT的张锋、国内包括北大在内的多个大学和研究机构的科学家也做出了贡献,特别是在改进、推广等方面。CRISPR/Cas9技术很快就被生物领域的众多实验室有效地用于多种生物,其简单而高效,一般生物学研究生都能很快上手,而且成为培训本科生的实验方法之一。
以前的方法只能在很少的生物中应用,所以不用担心,只有比较专业的人才能用,也减轻了人们的担忧,但CRISPR/Cas9的有效性和便利性带来了极大的冲击。
遗传和扩散:人类基因编辑
长期以来,科学界知道新的科学技术不能用于人类生殖细胞,特别是不能产出婴儿。因为历史和发展的原因,中国科学和技术方面的立法相对滞后,对于基因编辑依法管理是弱项。CRISPR/Cas9基因编辑方法的简易引起了前所未有的问题。
2015年,广州中山大学副教授黄军就由一个医院的委员会同意后,对人类生殖细胞进行了基因编辑(Liang et al。, 2015)。他们显然不清楚国际科学界的顾虑,而把文章当成突破寄给顶级杂志(英国的Nature、美国的 Science),被杂志编辑部因伦理问题而拒稿。而中国出版和主编的英文杂志《蛋白质和细胞》却在一天之内就快速收稿,决定发表。这一工作当即引起国际科学界很大的关注。值得一提的是,黄军就的工作选择了不能完成发育的异常胚胎,所以不会产生婴儿。在国际反应很强的情况下,中国有关部门并没有采取相应的举措。
2017年,前苏联移民美国的科学家Mitalipov 在Nature发表论文,报道他们编辑了人类生殖细胞的基因,其第一作者是华人(Ma et al。, 2017)。显然,当年拒绝黄军就论文的Nature现在不以生殖细胞基因编辑作为伦理的担忧。
2018年11月26日,南方科技大学停薪留职、在外开公司的贺建奎对媒体宣布,他不仅对人类生殖细胞进行了基因编辑,而且产出了一对婴儿。消息传出,有些国内主流中文媒体作为科技突破来发表消息,但国内外科学界立即哗然,国际媒体也纷纷质疑。
因基因编辑技术门槛低,现在有很多人可以做,如果不及时制止,就有泛滥的可能性。在很多年轻的学者来看,如果对现有情况不处理而后也不出台严格的法律法规,他们中有些人也可能会做,因为早在贺建奎之前,他们就在动物中做过,他们比贺建奎更加熟悉的基因编辑技术。
体细胞基因编辑不带来新的伦理问题
在人之外的动植物使用基因编辑技术,不会带来新的伦理问题,科学界和人类已熟悉、并且已有合适的审批和监管措施。

人类胚胎基因编辑的核心问题
在人类身上应用基因编辑技术,也要区分用是人的生殖细胞还是体细胞。成人的生殖细胞是指精子或卵子,其他细胞则为体细胞。在胚胎发育后期,也有明确的精子、卵子,而早期首先是精子和卵子形成的受精卵。受精卵含以后称为生殖细胞部分,而随着受精卵分裂,胚胎不断发育,胚胎部分细胞明确成为体细胞,部分成为生殖细胞的前体,以后成为生殖细胞。在生殖细胞和体细胞的来源没分开之前,我们需要考虑其生殖细胞的问题。在生殖细胞和体细胞分开后,我们的顾虑是生殖细胞,而不用担心体细胞。

人类生殖细胞基因编辑,是此事件的核心问题
体细胞为什么不需要格外担心?因为对于体细胞进行基因编辑,与现在我们大家接受的各种治疗方式一样,只影响个人,一般来说不会影响生殖细胞。体细胞基因编辑的伦理也就可遵循已有的规范,在了解科学可行和医学安全后,比较效果和副作用,也就是对个体的利弊,可由个人和家庭做出决定是否进行体细胞的基因编辑。
在长达三十多年的坚持之后,近年国际上在体细胞的基因治疗取得了很大的进展。对人的体细胞进行基因操纵,终于可治疗一些疾病,而且其发展趋势很好,有可能可以治疗越来越多的疾病。体细胞的基因治疗,应该得到积极的支持,使之越来越多地造福患者和社会。
人类生殖细胞的基因编辑缺乏临床必要性
对于遗传疾病来说,绝大多数无需生殖细胞基因编辑,而是通过选择自然产生的受精卵就可以做到。
北京大学化学家谢晓亮和北大三院妇产科专家乔杰合作的研究,就可有效地检测受精卵的基因型,从而选择合适的进行植入(Yan et al。, 2015)。对一般的基因变化造成的疾病罹患性,基因测序后可选择理想的受精卵植入子宫受孕,就可以解决。对于罕见的父母都是纯合子的基因变化,可通过他们在结婚前自愿披露自己的状况而避免相互结婚,如果他们不披露或者知道双方情况还要结婚,是自愿承担带来的患病代价。
基因编辑在技术上优于选择自然的受精卵,是对同时得到多个理想的基因和基因型,以及将个人基因组不存在的基因型编辑到后代中。后两点都带来极大的伦理问题,应该由国家立法进行规范。
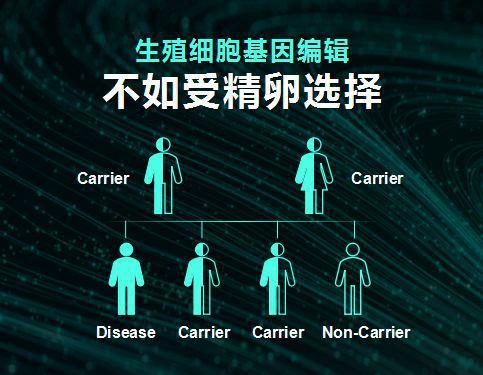
生殖细胞的基因编辑不如受精卵选择
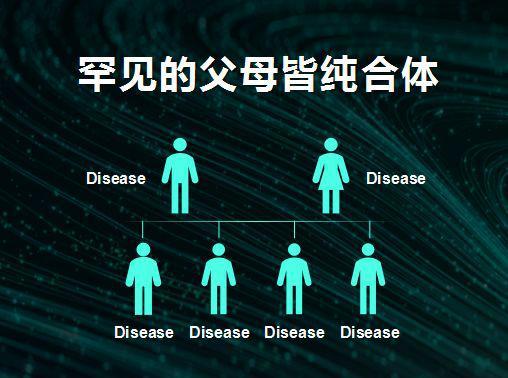
罕见的父母皆为纯合体可通过婚前自愿披露遗传信息,减少后代患病的可能性
人的多数基因(分布在常染色体的基因)有两个拷贝,一个在来自父亲的染色体,一个在来自母亲的染色体。如果用+代表正常,用*代表致病突变,那么一个人的基因型就可能有三种:+/+,+/*;*/*。+/+不会导致疾病,大多数情况下*/*,少数情况+/*以及*/*基因型都能导致疾病。一个婴儿的父亲或母亲一般很少同时带有同一个致病的突变基因,而如果出现父亲是+/*,母亲也是+/*,那么他们怀孕的受精卵四分之一的概率将是+/+,四分之二是+/*,剩下的四分之一则是*/*。对于大多数疾病来说,只有*/*会致病,而其他两种情况(+/+和+/*)不会致病,那么在体外受精(IVF)时选择用这两种就可以,无需基因编辑。即使+/*会致病,也可以选择使用+/+的受精卵。这在技术上可行的,只需要通过选择不会致病的受精卵即可,不会带来新的伦理问题。最难办的是,父母都是*/*基因型(或显性遗传时父母一方为*/*),那么他们的后代都将患病。但这些情况非常罕见,人群中一般+/+最多,+/*次之,父母都是+/*就很少,而父母都是*/*的情形则极为罕见。国家的规范应该确定是否允许这种父母进行生殖细胞基因编辑,还是允许个人进行基因检测得知自身携带*/*,结婚前自愿披露自己的基因型。这是一个两难的问题:是A(允许*/*的父母进行生殖细胞基因编辑),还是B(允许披露自己或知道对方的基因型)。事实上,B的代价远远小于A,B影响的是极少的人,不影响人类。由此我们不能因为有罕见的情况,而允许人类普遍进行生殖细胞的基因编辑。
对自然产生的人类生殖细胞进行测序后加以选择还是基因编辑?第一个差别在于,选择的基因有限,对于一个受精卵来说,如果只有一个或少数几个疾病罹患基因,选择是容易的,但如果需要对多个基因进行选择,很可能不能选到同时多个基因都理想的。基因编辑技术在理论上可对多个基因进行编辑,虽然目前还不是特别容易,但今后可能做到。但第二个差别在于,推动人类生殖细胞的基因编辑,实际很可能希望不限于疾病,而是其他性状,如肌肉更多、运动和其他能力提高,这样才有非常广泛的市场,“值得”他们的商业利益。
所以人类生殖细胞基因编辑的问题,实际是商业推广大规模基因编辑与人类对于群体基因分布安全性的问题,对这一问题带来的风险远高于少数人因为一个基因被编辑而出现的个人安全性风险。
人类生殖细胞基因编辑带来的安全危机
为什么需要格外担心生殖细胞的基因编辑?这不是人文关怀,而是严谨的科学问题。

人类胚胎的基因编辑带来的危害
1) 个体选择不能强加于人类社会。因为生殖细胞的基因编辑后,被编辑的个人会和其他人婚配,其后代也会带有编辑后的基因。因此,生殖细胞基因编辑的影响就不限于个人,而会流入人群。生殖细胞基因编辑就会影响群体、国家和人类社会。因此,是否进行个人的生殖细胞基因编辑,就不仅仅是个人和家庭的事情,而是有关群体、国家和人类社会的事情。
2) 一旦开启人类生殖细胞基因编辑则无终点。概念上,无法分开疾病和非疾病。数量上,无法限定一次只能编辑一个基因还是多个基因。具体不能限定哪些基因可以动、哪些不可以动。也就可能出现同时大规模变化很多基因,而且各行其是,出现很大的混乱,如一个人或一个家庭希望不患艾滋病,另外一个人或一个家庭希望治疗乳腺癌,还有家庭希望肌肉多一些、个子高一些……目前,这些都有已知的基因,有已知的突变,如myostatin基因无论在老鼠、猪、牛还是人,如果缺失掉,可让肌肉增多。不同的人和家庭都可以认为自己有道理,并认为应该限制其他人提议的编辑而允许自己希望的基因编辑:你可以预防癌症,我家好几代肌肉少吃了亏,为什么不能增加肌肉;越来越多的研究发现影响学习记忆、影响运动成绩的基因变化,人类是否需要对生殖细胞进行基因编辑,有利于他们的后代成为学者或运动员;某个时代时髦什么,就往那个方向编辑,还是怎么投票怎么编辑;如果因为今天Trump“成功”了,很多人要模仿他的基因,这怎么办;当年可是有很多德国人给孩子取名阿道夫(Adolf)效仿阿道夫·希特勒,名字可换来换去,生殖细胞的基因编辑至少要在后代待一代,有人肯定能承受。
3) 能否限制基因编辑只用于改变疾病相关的基因变化,而不用于增强功能?这种想法是不了解一个基本事实:疾病是人们主观定义的。例如,精神疾病目前全世界是用美国精神病学会的诊断书,这本书过几年有其委员会修订一次,有时出现新的疾病定义,有时删除以前的疾病定义。又比如有些人说自己长期疼痛,而其他人无法知道他/她是否痛。即使有客观指标的疾病,对生殖细胞的基因治疗也有代价,如镰刀状红细胞贫血是基因变化造成,基因编辑修复突变可改善贫血但更容易得疟疾。北欧有一家族,多代获得了滑雪冠军,因为他们家遗传一个基因突变(红细胞生成素受体EPOR的基因突变)。如果有一些人自己就能决定后代基因变成这样,那么滑雪冠军(和其他需要红细胞量的运动员)将会增加,但会带来血管容易被堵住的相关疾病。其他基因变化,我们有时只知道带来的问题,不知道带来的好处,只表明我们目前知道的范围,不表明我们能够排除进化过程已经筛选过,有其好处。
4) 因为技术的简便,如果放开做人类的生殖细胞基因编辑,可能出现一个家庭希望改变一个基因,另外一个家庭希望改变另外一个基因,很多家庭希望改变很多基因。在有限知识的情况下,开展编辑基因竞赛,可能导致对人类的灾难。
5) 包括人类在内的生物,其基因需要一定程度的多样性,这是进化的常识。人类在进化过程中逐渐建立的基因库,人的所有基因都参与某种功能,没有用的基因在进化中会被淘汰,剩下都是有用的,而且自然的基因变化保留了对很多问题的防范。但科学家并不清楚到底多样性在什么范围,无法回答哪些可以减少一些多样性,哪些不可以。如果因为生殖细胞基因编辑按照一时一地的想法和便利,突然地大规模的人为改变,可能导致人类难以维持进化选择合适的基因库,从而危及人类,如在有关脑的基因变化中,我们目前不清楚所谓罹患精神病的基因变化,是否也会让脑进化得更聪明,更有创造力的基因变化,如果为了避免少数人患痴呆或精神病,把人类的脑进化停止在今天,整个人类是否愿意,当外界出现危机时,人类头脑有没有足够的应付能力?

人类生殖细胞基因编辑的推动者
6) 人类生殖细胞基因编辑的推动者,虽然现在打着为解决父母双方都携带基因变异纯合体(父母的两套染色体都坏了,也就是总共四条染色体坏了)的问题,其实因为这种情况极为罕见,推动者并不能获得商业利益,所以他们的目的是以这种借口说服突破对于生殖细胞基因编辑的禁止,从而进一步推动影响人类其他非疾病性状的基因编辑,他们还会推动对于一个受精卵的多个基因编辑。因为无法达成社会共识可编辑哪些基因,只要允许人类生殖细胞基因编辑,很难避免出现混乱的情况;
7) 生殖细胞细胞基因编辑,还有技术的安全性问题。例如脱靶问题,目的是编辑A基因,结果还有其他基因被改变了,其中有些改变可能带来安全问题。类似的技术问题已被广泛讨论。基因编辑对个人安全虽然重要,但其重要性低于以上对人类的影响。解决了技术对个人的安全性问题,并非就可以施行生殖细胞的基因编辑,还有本文上面讨论的其他问题。
国际问题:不能盲目跟风,而是加以甄别
在国际上,人类生殖细胞和胚胎有清晰的红线。其中胚胎的红线不仅是科学而且有宗教原因。但仅仅从科学上,就画出红线,严禁人工改造生殖细胞后植入子宫继续发育成婴儿。
中国有关部委曾经颁布一些规定。如1993年,卫生部药政局制定了《人的体细胞基因治疗临床研究质控要点》,当时因为技术远没有达到对人类生殖细胞基因编辑,所以没有规范生殖细胞的部分;2003年,科技部和卫生部发布的《人类胚胎干细胞研究伦理指导原则》虽然针对干细胞,也可以引申到基因编辑的生殖细胞,其第六条“利用体外受精、体细胞核移植、单性复制技术或遗传修饰获得的囊胚,其体外培养期限自受精或核移植开始不得超过14天。不得将前款中获得的已用于研究的人囊胚植入人或任何其他动物的生殖系统”。
法国、德国、英国等国的法律禁止人类生殖细胞进行基因改造,违法有刑事责任。美国只禁止用国家经费进行人类生殖细胞基因编辑的研究和应用,允许使用私人经费进行研究,但不支持编辑后植入子宫完成发育。
对于外国文化和科学家,中国需要避免国际上不适用于中国的两极问题。一极是基督教等宗教认为人体不可更改,人不能做上帝,否定理性和科学;一极是有些人(包括知名科学家)有商业利益。2017年,美国国家科学院组织人类基因编辑伦理会议,由于与会的积极支持者占多数,中立者很少,反对者则很少被请去参加会议,缺乏所谓“国际”代表性,有些是单独邀请美国之外的少数个人,而后者明确不代表所在国家。另外,西方讨论时明确提出立法可能失去与中国的竞争。所以美国科学院现有的会议需要参考,但不能简单照搬。哈佛医学院院长George Daley背景局限,焦点只在疾病,对遗传学不够熟悉,不谈疾病有没有基因编辑的刚需,不谈一旦开放人类生殖细胞的基编辑,就难以阻止对影响非疾病人类性状的多个基因进行大规模编辑的可能性。Daley的观点也被包括现任NIH院长Francis Collins在内的遗传学家批评。
有美国科学家,打着为罕见的父母双方都为纯合体家庭的利益考虑的旗号,而坚持要推进生殖细胞基因修饰,其实他们自己开了公司,他们的公司不可能通过帮助罕见的纯合体家庭而获利,其目的是为了生殖细胞基因修饰增强功能,在科学内行看就是“司马昭之心”,但专业之外的人可能被其美国大学的名气,科学贡献所迷惑,认为不可能出错,不可能忽悠,这时我们要记住的一个反例是Trump就获得很多美国人支持的。
有些美国科学家,在本国禁止或不支持的情况下,怂恿和帮助中国的科学家做生殖细胞基因编辑,然后转身对美国说:看中国都在做了,我们不能禁止,我们不能竞争失败,从而抛弃伦理的辩论,绕开道德和公众监督,利用中国达到其商业目的。我们应该对美国科学家的意见持谨慎态度,分清其公德公益和私德私利。
在一定时间、空间范围内,获得有利于社会、国家和人类的社会共识并非易事。特别是在科学、技术等需要一定知识背景,且不被哗众取宠者通过情绪喧闹而影响政策法律法规的制定。西方曾经对公众调查对基因编辑的接受性,多数人接受(Scheufele et al。, 2017),但这种调查有意无意不区分体细胞和生殖细胞基因编辑,且公众是否大部分知道体细胞和生殖细胞的差别恐怕也有很大的疑问。最近由CRISPR/Cas9发明者之一Doudna写的科普书,居然完全没有讨论生殖细胞基因编辑的问题,而只简单提倡基因编辑可用于人类(Doudna and Sternberg, 2017)。
解决方法:亟需立法,建立国家咨询机构

中国的胚胎基因编辑解决方法
1) 对于基因编辑,急需亡羊补牢,由国家建规立法,明确地、严格地规范对基因编辑,允许监管下的体细胞基因编辑,禁止人类生殖细胞基因编辑;
2) 建立健全的制度,审核、批准都有质量过硬、敢于担当的机构。加强执法,对违反规章制度、违反法律法规的个人和单位严惩不怠;
3) 对于日新月异的科学领域,需要建立国家咨询机构,经常、及时交流促进国家保持对科技发展的敏感性,以便今后不出现或少出现因为科技发展而惊世骇俗地突破底线的问题,更要预防危害人类社会安全的问题;
4) 积极建立国际交流合作,协调重大问题,推动建立国际公约,并转化为各国建立全人类利益一致的法律法规。
注:1月15日,“知识分子”、“赛先生”以及中关村海淀园管委会联合主办了“科学精神中国行”新年专场活动,本文整理于生命科学家、北京大学教授饶毅在活动中发表的主旨演讲“基因编辑与人类未来”。
参考文献
Avery OT, MacLeod CM and McCarty M (1944)。 Studies on the chemical nature of the substance inducing transformation of pneumococcal types.Journal of Experimental Medicine 79: 137-158。
Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero DA, Horvath P(2007) CRISPR provides acquired resistance against viruses in prokaryotes。 Science 315:1709–1712。
Bolotin A, Quinquis B, Sorokin A, Ehrlich DS (2005) Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin。 Microbiology 151:2551–2561。
Brouns SJ, Jore MM, Lundgren M, Westra ER, Slijkhuis RJH, Snijders APL, Dickman MJ,Makarova KS, Koonin EV, and van der Oost J (2008) Small CRISPR RNAs guideantiviral defense in prokaryotes。 Science321:960–964。
Cohen SN, Chang ACY, Boyer HW and Helling RB (1973)Construction of biological bacterial plasmids in vitro。 Proceedings of the National Academy of Sciences USA 70:3240-3244。
Deltcheva E, Chylinski K, Sharma CM, Gonzales K, Chao Y, Pirzada PA, Eckert MR, Vogel Jand Charpentier E (2011) CRISPR RNA maturation by trans-encoded small RNA andhost factor RNase III。 Nature471:602–607。
Deveau H, Barrangou R, Garneau JE, Labonté J, Fremaux C, Boyaval P, Romero DA, Horvath P, Moineau S (2008) Phage response to CRISPR-encoded resistance in Streptococcus thermophilus。 Journal of Bacteriology 190:1390–1400。
Doudna JA and Sternberg, SH (2017) A crack in creation。 Gene editing and the unthinkable power to control evolution。 Houghton Mifflin Harcourt, Boston。
GarneauJE, Marie-Ève Dupuis, Villion M, Romero DA, Barrangou R, Boyaval P, Fremaux C, Horvath P, Magadán AF and Moineau S (2010) The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA。 Nature 468:67–71。
Groenen PM, Bunschoten AE, van Soolingen D and van Embden JDA (1993) Nature of DNApoly morphism in the direct repeat cluster of Mycobacterium tuberculosis; application for strain differentiation by a novel typing method。 Molecular Microbiology 10:1057–1065。
Haft DH, Selengut J, Mongodin EF and Nelson KE (2005) A guild of 45 CRISPR-associated (Cas) protein families and multiple CRISPR/Cas subtypes exist in prokaryotic genomes。 PLoS Computational Biology 1:e60。
Hale CR, Zhao P, Olson S, Duff MO, Graveley BR, Wells L, Terns RM and Terns MP(2009) RNA-Guided RNA cleavage by a CRISPR RNA-Cas protein complex。 Cell 139:945–956。
Hale CR, Majumdar S, Elmore J, Pfister N, Compton M, Olson S Resch AM, Gover IIIC VC, Graveley BR, Terns RM and Terns MP (2012) Essential features and rational design of CRISPR RNAs that function with the Cas RAMP module complex to cleave RNAs。 Molecular Cell 45:292–302。
Hermans PW, van Soolingen D, Bik EM, de Haas PEW, Dale JW, and van EMbden JDA (1991)Insertion element IS987 from Mycobacterium bovis BCG is located in a hot-spot integration region for insertion elements in Mycobacterium tuberculosis complex strains。 Infection and Immunity 59:2695–2705。
Horvath P and Barrangou R (2010) CRISPR/CAS, the immune system of bacteria and archaea.Science 327:167–170。
Horvath P, Romero DA, Coûté-Monvoisin A-C, Richards M, Deveau H, Moineau S, Boyaval P, Fremaux C and Barrangou R (2008) Diversity, activity, and evolution of CRISPR loci in Streptococcus thermophilus。 Journal of Bacteriology 190:1401–1412。
Ishino Y, Shinagawa H, Makino K, Amemura M, and Nakata A (1987) Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion inEscherichia coli, and identification of the gene product。 Journal of Bacteriology 169:5429–5433。
Jansen R, van Embden JDA, Gaastra W, Schouls LM (2002) Identification of genes that are associated with DNA repeats in prokaryotes。 Molecular Microbiology 43:1565–1575。
Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA and Charpentier E (2012) A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity。 Science337:816-827。
Liang R, Xu Y, Zhang X, Ding C, Huang R, Zhang Z, Lv J, XieX, Chen Y, Li Y, Sun Y, Bai Y, Songyang Z, Ma W, Zhou C and Huang J (2015)CRISPR/Cas9-mediated gene editing in human tripronuclear zygotes。 Protein and Cell 6:363-372。
Ma H, Marti-Gutieerez N, Parks SW, Wu J, Lee Y, Suzuki K,Koski A, Ji D, Hayama T, Ahmed R, Darby H, Van Dyken C, Li Y, Kang E, Park AR,Kim D, Kim ST, Gong J, Gu Y, Xu X, Battaglia D, Krieg SA, Lee, DM, Wu DH, WolfDP, Heitner SB, Belmonte, JCI, Amato P, Kim JS, Kaul S, Mitalipov S (2017)Correction of a pathogenic gene mutation in human embryos。 Nature 548:413-419。
Makarova KS, Grishin NV, Shabalina SA, Wolf YI and Koonin EV (2006) A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action。 Biology Direct 1:7。
Marraffini LA and Sontheimer EJ (2008) CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA。 Science322:1843–1845。
Maxam A and Gilbert W (1977) A new method for sequencing DNA。 Proceedings of the National Academy of Sciences USA 74:560–564。
Miescher JF (1871) Ueber die chemische Zusammensetzung der Eiterzellen。 Medisch-chemische Untersuchungen4:441-460。
Mojica FJM, Juez G and Rodríquez-Valera F (1993) Transcription at different salinitiesof Haloferax mediterranei sequences adjacent to partially modified PstI sites。 Molecular Microbiology 9:613–621。
Mojica FJ, Ferrer C, Juez G and Rodríquez-Valera F (1995) Long stretches of short tandem repeats are present in the largest replicons of the Archaea Haloferax mediterranei and Haloferax volcanii and could be involved in replicon partitioning。 Molecular Microbiology17:85–93。
Mojica FJ, Díez-Villaseñor C, Soria E and Juez G (2000) Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria。 Molecular Microbiology36:244–246。
Mojica FJ, Díez-Villaseñor C, García-Martínez J and Soria E (2005) Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements。 Journal of Molecular Evolution60:174–182。
Mojica FJ, Díez-Villaseñor C, García-Martínez J and Almendros C (2009) Short motif sequences determine the targets of the prokaryotic CRISPR defense system。 Microbiology 155:733–740。
Nakata A, Amemura M and Makino K (1989) Unusual nucleotide arrangement with repeated sequences in the Escherichia coli K-12 chromosome。 Journal of Bacteriology 171:3553–3556。
National Academy of Sciences and National Academy of Medicine, Human Genome Editing: Science, Ethics, and Governance (National Academies Press, Washington, DC, 2017)
Pourcel C, Salvignol G and Vergnaud G (2005) CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies。 Microbiology151:653–663。
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors。 Proceedings of the National Academy of Sciences USA 74:5463–5467。
Scheufele DA, Xenos MA, Howell EL, Rose KM, Brossard D and Hardy BW (2017) US attitudes on human genome editing。 Science 357:553-554。
Thomas KR and Capecchi MR (1987) Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells。 Cell 51:503–512。
Watson JD and Crick FHC (1953)。 A structure for deoxyribose nucleic acid。 Nature 171:737-738。
Yan L, Huang L, Xu L, Huang J, Ma F, Zhu X, Tang Y, Liu M,Lian Y, Liu P, Li R, Lu S, Tang F, Qiao J and Xie XS (2015) Live births after simultaneous avoidance of monogenic diseases and chromosome abnormality by next-generation sequencing with linkage analyses。 Proceedings of the National Academy of Sciences USA 11:15964–15969。
Zeng Y, Li J, Li G, Huang S, Yu W, Zhang Y, Chen D, Chen J, Liu J and Huang X (2018)Correction of the Marfan syndrome pathogenic FBN1 mutation by base editing in human cells and heterozygous embryos。 Molecular Therapy 26:2631-2637。
推荐
《科学大家》专栏投稿邮箱:sciencetougao@sina.com 来稿请注明姓名、单位、职务


