


在许多物种中,雄性会发育出明显的性特征来吸引雌性,并通过竞争来排挤潜在的雄性对手。最具代表意义的例子包括雄性孔雀或极乐鸟那夸张、艳丽的羽毛。
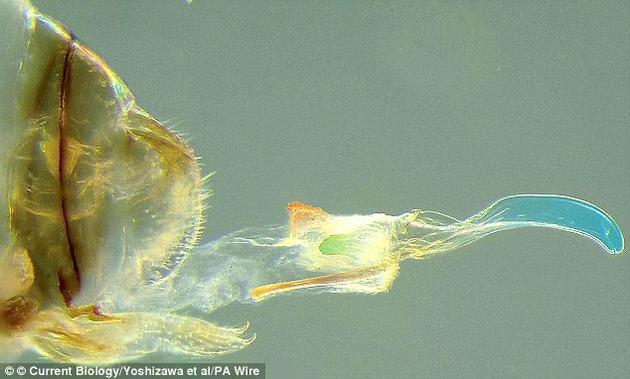
在一类属名为Neotrogla(啮虫目小啮虫亚目下的一个属,如图)的洞穴昆虫中,雄性没有插入式的、类似阴茎的结构,而雌性则发育出了类似阴茎的结构(称为gynosoma)
新浪科技讯 北京时间3月2日消息,在许多物种中,雄性会发育出明显的性特征来吸引雌性,并通过竞争来排挤潜在的雄性对手。最具代表性的例子包括雄性孔雀或极乐鸟那夸张、艳丽的羽毛,以及欧洲马鹿群体中,占统治地位的雄性所具有的巨大鹿角。但是,自然界中不同性别的角色是如何确定的?为什么雄性通常都要通过竞争才能接近雌性?
研究者认为,这些问题的答案就在于所谓的“贝特曼原理”(Bateman principle)。该原理指出,对于在后代身上投资更少的性别,性选择的作用更加强烈。
1972年,美国演化生物学家罗伯特·泰弗士(Robert Trivers)提出了“亲代投资”的概念,认为这是决定何种性别处于更高性选择压力的关键因素。根据泰弗士的理论和贝特曼原理,对于将较少资源分配到亲代投资的性别,其所受到的性选择压力就更大。
产生精细胞的成本要比产生卵细胞的成本低得多。这意味着,雄性能产生的配子——在有性繁殖中结合的细胞——数量比雌性多得多。反过来,两性因此受到的影响也非常不同。通常情况下,雌性会通过孵卵、哺育和保护等行为,对后代进行更多的投资。

一些物种,比如某些种类的豆娘(如图),雄性甚至会把此前其他雄性留在雌性体内的精子清除掉。

雄性海马负责孵化、照料受精卵,在幼鱼能够自由活动时再将其释放出育儿袋。
因此,雌性的亲代投资比雄性要大得多,而雄性需要经过竞争才能接近雌性。然而,自然界中还存在不少性别角色倒转的例子,其中雌性需要通过比雄性强度更大的竞争,才能获得交配的机会。这些例子包括雉鸻科水禽、新西兰的几维鸟、原产于中美洲和南美洲的䳍形目鸟类,以及一些涉禽物种。
此外,还得提到海马。雄性海马负责孵化、照料受精卵,在幼鱼能够自由活动时再将其释放出育儿袋;两栖动物中,一些青蛙和箭毒蛙属物种也有类似行为;哺乳动物中,性别角色转换的例子有转角牛羚(学名:Damaliscus lunatus)。
与性角色转变有关的生理变化中,最为令人称奇的过程出现于某些鱼类,如雌雄同体的金头鲷(学名:Sparus aurata)。在鱼卵孵化时,所有的金头鲷个体都是雄性,而当它们长到一定年龄,就会根据重量、荷尔蒙和群体因素等条件的不同,而选择是否变成雌性。
在有性繁殖的动物中,性器官的形态一直是最多样而且演化最快的特征之一。在传统性别角色的物种中,由于高强度竞争引起的强性选择会导致雄性出现比雌性更加复杂的性器官。雄性通常表现出生殖器或射精(包括精液中的蛋白质)等方面的适应特征,从而更快地输送精子,刺激雌性更快接收精子,或将其他同性对手竞争下去。一些物种,比如某些种类的豆娘,雄性甚至会把此前其他雄性留在雌性体内的精子清除掉。

与性角色转变有关的生理变化中,最为令人称奇的过程出现于某些鱼类,如雌雄同体的金头鲷(学名:Sparus aurata)。

阴蒂结构会从雌性斑鬣狗(如图)的身体向外突出十几厘米,而且开口非常狭窄,这使雄性很难与之进行交配,在生出幼崽时也非常困难,头胎死亡率非常高。
在哺乳动物中,雌性(学名:Crocuta crocuta)甚至发育出了“假阴茎”的结构——延长的阴蒂,这是由于幼崽发育最后阶段时荷尔蒙快速分泌导致的结果。阴蒂结构会从雌性身体向外突出十几厘米,而且开口非常狭窄,这使雄性很难与之进行交配,在生出幼崽时也非常困难,头胎死亡率非常高。尽管从性器官的功能上看它们是雌性,但实际上,它们扮演着类似其他物种雄性的角色。
另一个不可思议的例子是一类属名为Neotrogla(啮虫目小啮虫亚目下的一个属)的洞穴昆虫。这类昆虫的雄性没有插入式、类似阴茎的结构,而雌性则发育出了类似阴茎的结构(称为gynosoma),可以伸入雄性体内采集所谓的精囊(spermatophore)。许多无脊椎动物和一些脊椎动物(如蝾螈)都会使用精囊作为保存精子的容器。在Neotrogla属昆虫的例子中,雌性利用充满创意且独一无二的似阴茎器官将精子从雄性体内拖出来。
这一器官还具有棘刺,使雌性能从内部与雄性固定在一起。是什么导致了这种结构的出现?答案很简单:对精液的性竞争和性冲突。Neotrogla属昆虫栖息在水和食物都很匮乏的洞穴中。在这样的条件下,争夺精子的竞争非常激烈,雌性要努力寻找交配的机会。
当一只雌性找到一只雄性时,其gynosoma结构能使其锚定在雄性身上,进行较长时间的交配。
自然界中存在着许多常见的交配方式,但也有少数的例外。这些例外使我们更加了解自然界中动物演化的机制,有性繁殖的方式并不像传统认为的那么老套。相反,不同性别的行为取决于多种因素,如亲代投资的不对称、性别比例或配偶的易得性等。因此,下次如果你看到一只性器官突出的动物,请不要想当然地认为它就是雄性。(任天)
